Deficiency of 15-LOX-1 Induces Radioresistance through Downregulation of MacroH2A2 in Colorectal Cancer

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
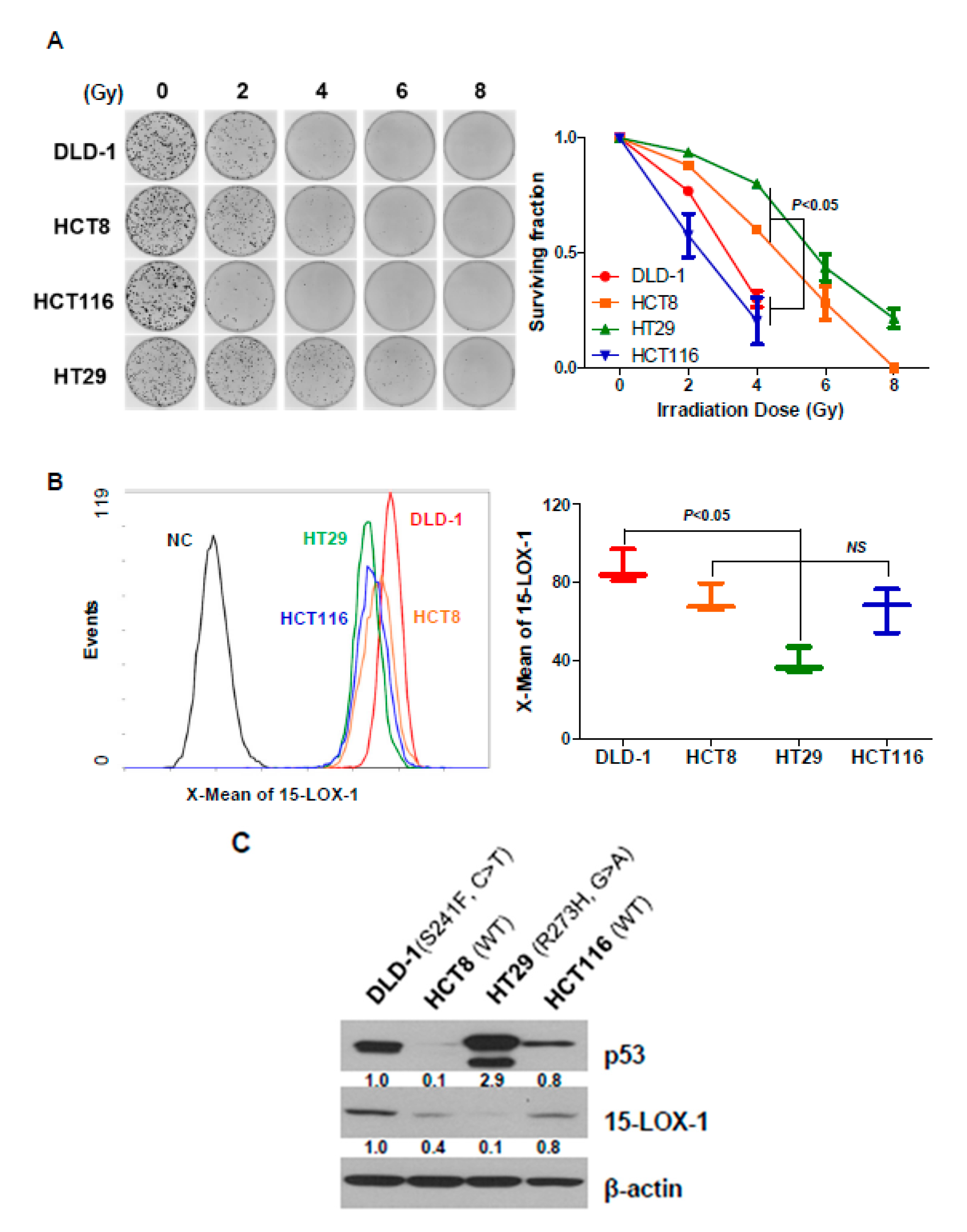
2.1. Radiosensitivity of CRC Cell Lines Correlates with 15-Lox-1 Expression Levels
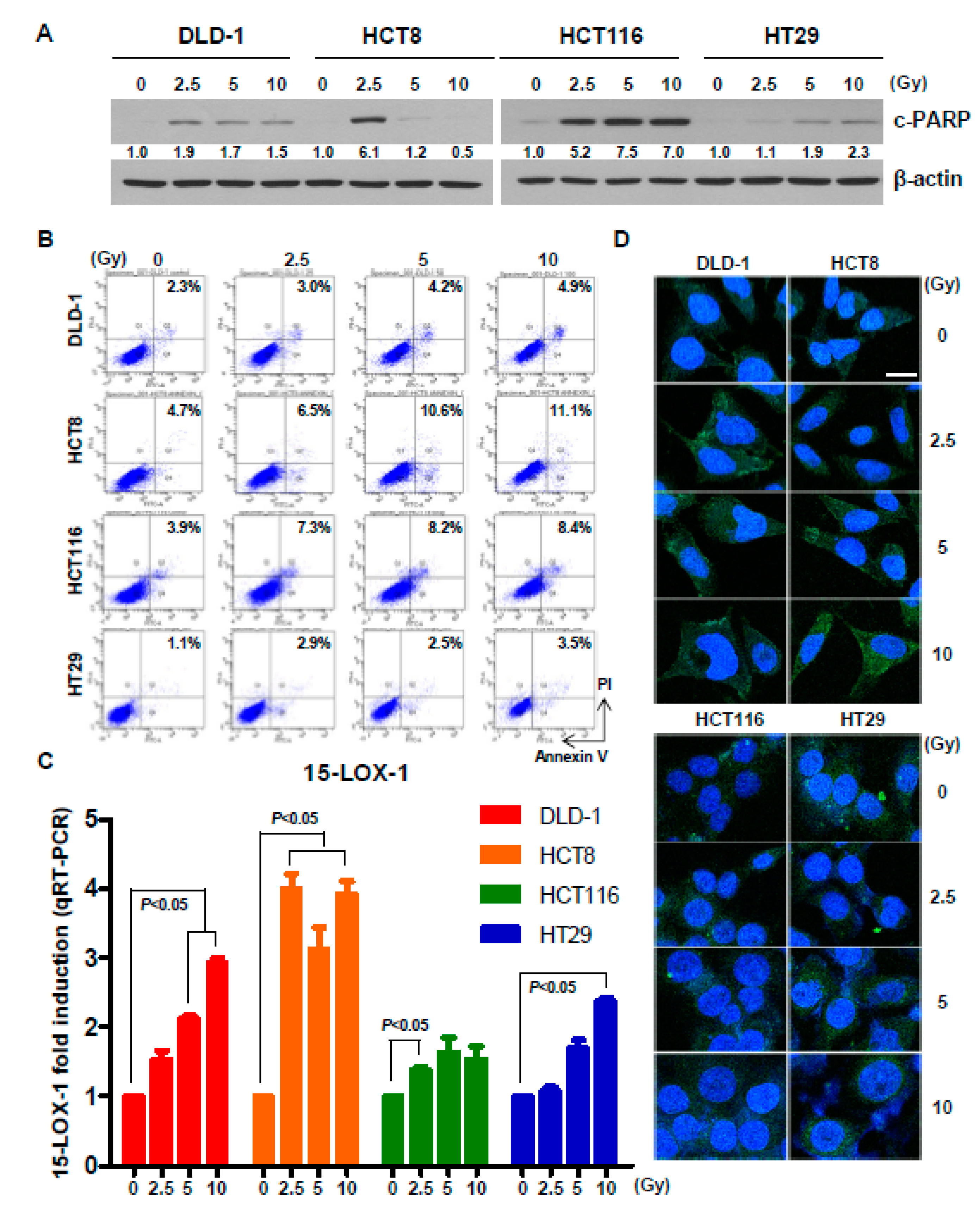
2.2. Radiation Induces Cell Death and Upregulates 15-Lox-1 Expression
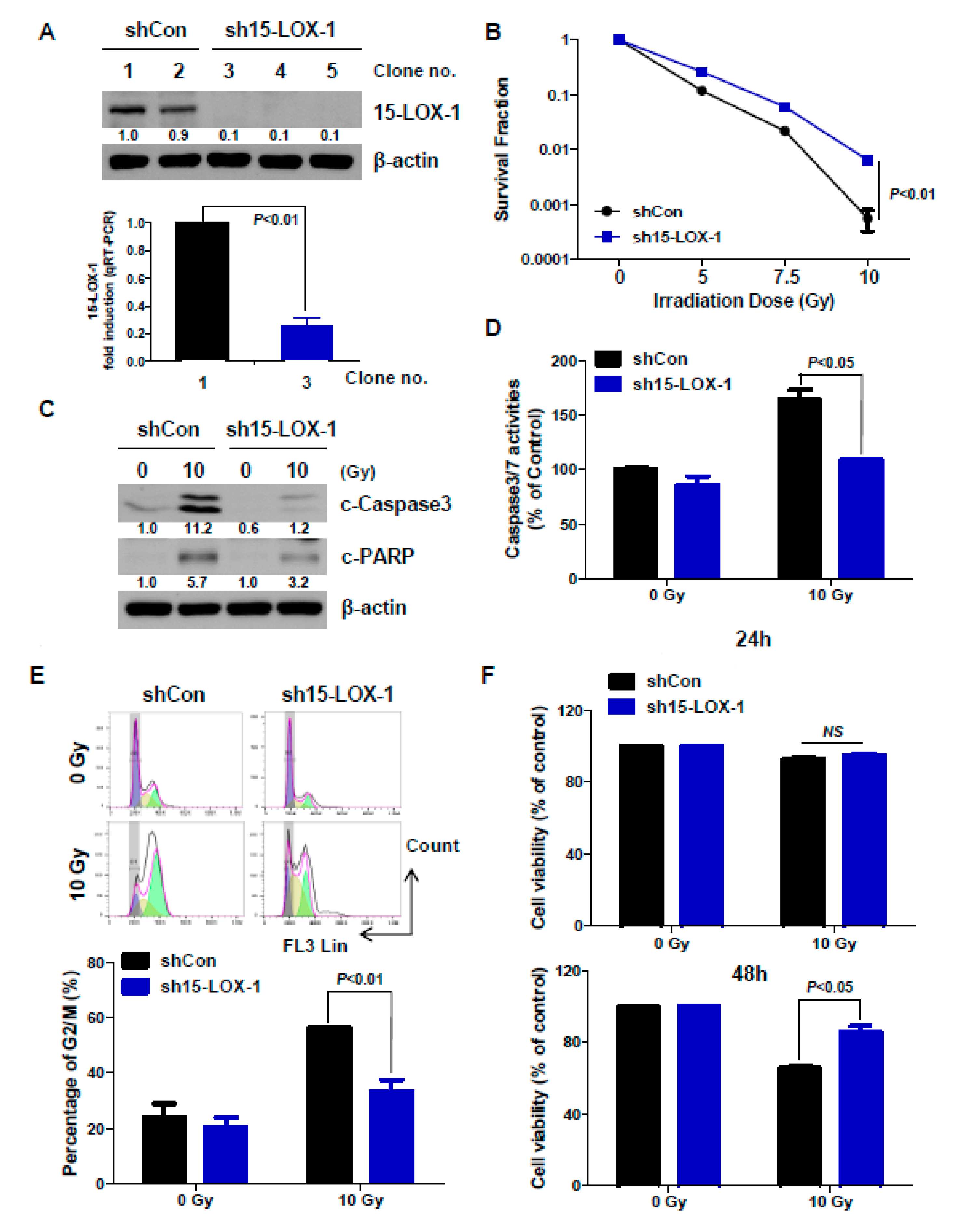
2.3. The Absence of 15-Lox-1 Decreases Radiation Sensitivity
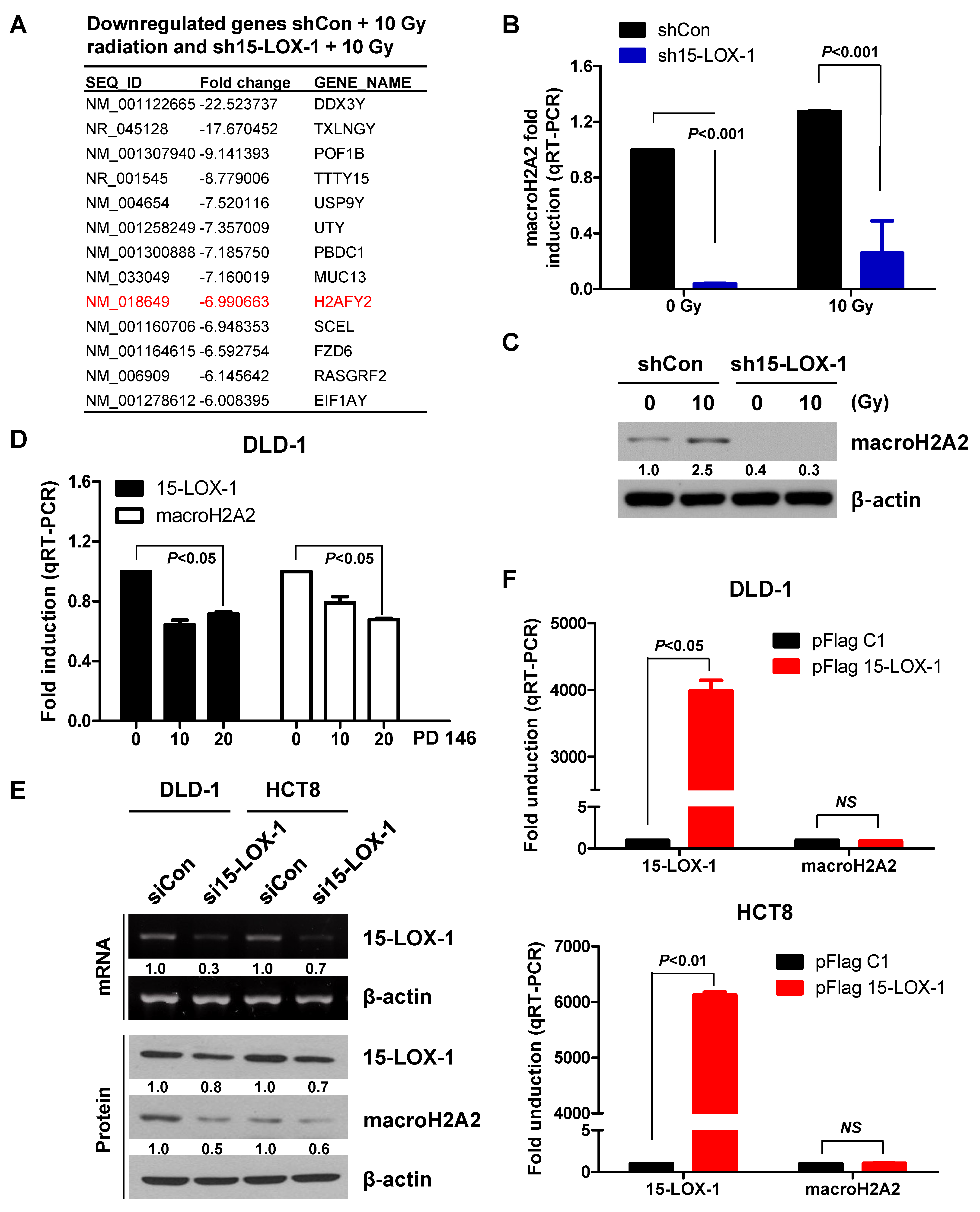
2.4. MacroH2A2 is Transcriptionally Regulated by 15-Lox-1 in Crc Cells
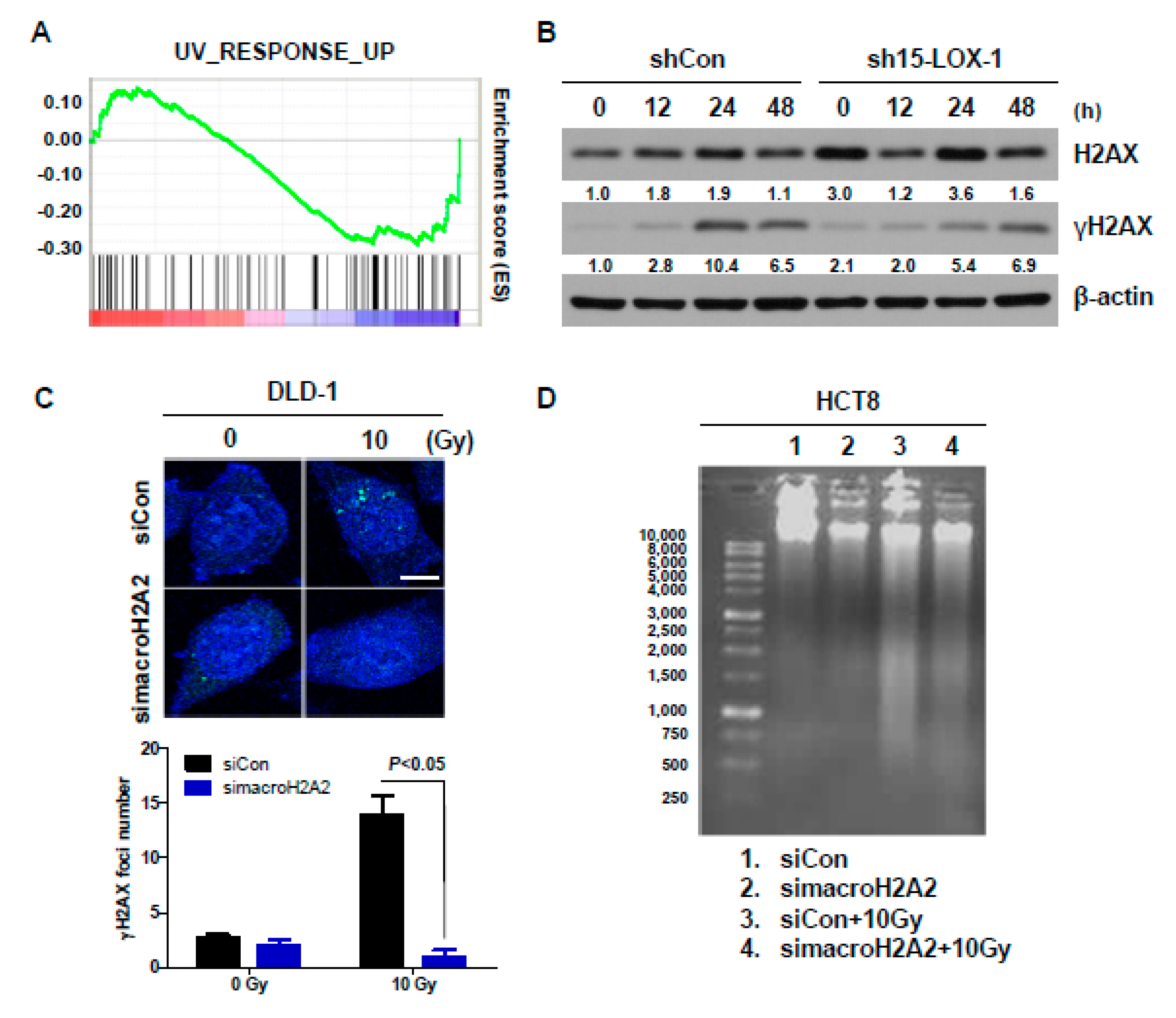
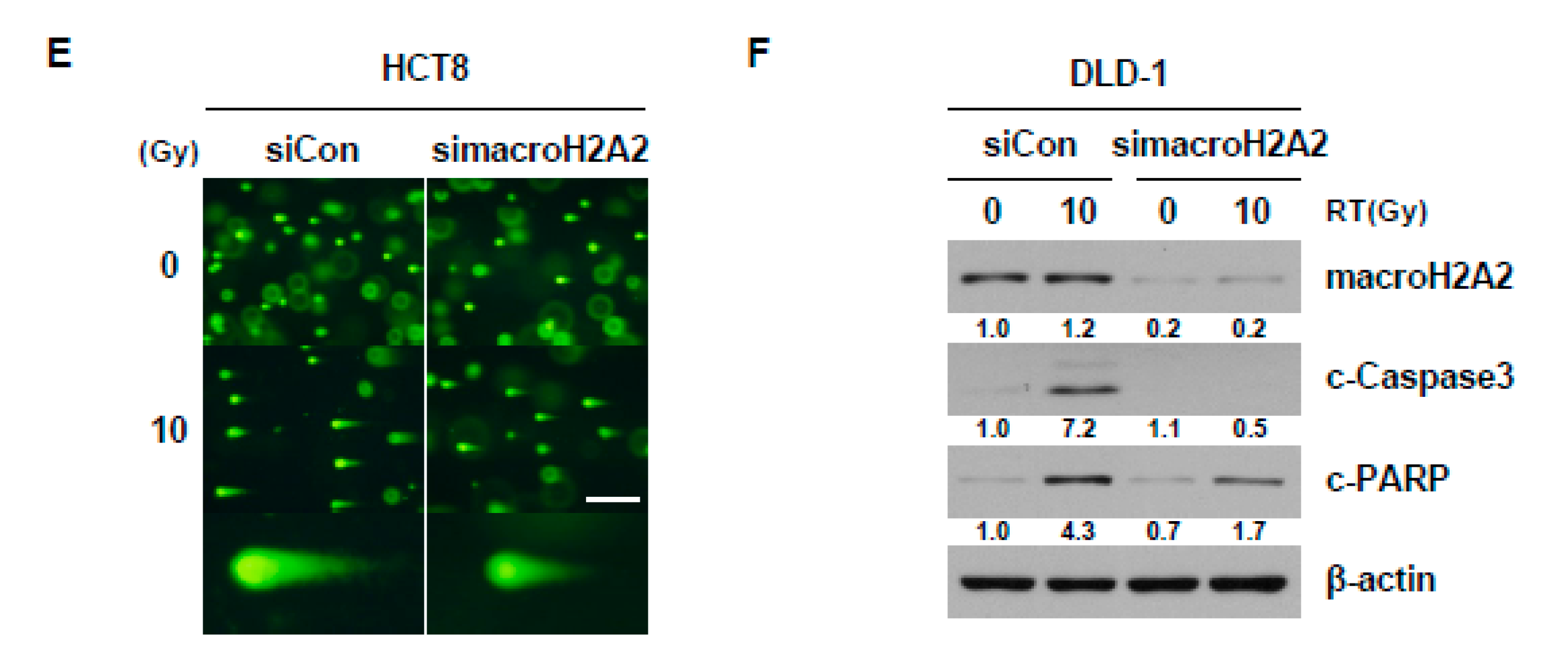
2.5. Reduction in macroH2A2 Function Caused by 15-Lox-1 Transcriptional Downregulation Is Involved in Radioresistance through Suppression of the Radiation Response
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Reagents and Antibodies
4.3. Western Blot Analysis
4.4. Polymerase Chain Reaction (PCR)
4.5. Clonogenic Cell Survival Assay
4.6. Microarray Analysis
4.7. Transfection
4.8. Cell-Cycle Assay
4.9. Analysis of 15-LOX-1 Using Flow Cytometry
4.10. Immunofluorescent Staining
4.11. Caspase 3/7 Assay
4.12. Annexin V Assay
4.13. Cell Viability Assay
4.14. Apoptotic DNA Ladder Detection
4.15. Comet Assay
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wu, J.; Chen, Y.; Geng, G.; Li, L.; Yin, P.; Nowsheen, S.; Li, Y.; Wu, C.; Liu, J.; Zhao, F.; et al. USP39 regulates DNA damage response and chemo-radiation resistance by deubiquitinating and stabilizing CHK2. Cancer Lett. 2019, 449, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Liu, L.; Liu, X.; Qi, L.; Wang, W.; Zhao, G.; Wang, L.; Dai, Y. Activation of PTGS2/NF-kappaB signaling pathway enhances radiation resistance of glioma. Cancer Med. 2019. [Google Scholar] [CrossRef] [PubMed]
- El Bezawy, R.; Tinelli, S.; Tortoreto, M.; Doldi, V.; Zuco, V.; Folini, M.; Stucchi, C.; Rancati, T.; Valdagni, R.; Gandellini, P.; et al. miR-205 enhances radiation sensitivity of prostate cancer cells by impairing DNA damage repair through PKCepsilon and ZEB1 inhibition. J. Exp. Clin. Cancer Res. 2019, 38, 51. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Liu, G.; Huang, K.; Zheng, Q.; Li, Y.; Yu, C. Hypoxia-Induced Upregulation of HE4 Is Responsible for Resistance to Radiation Therapy of Gastric Cancer. Mol. Ther. Oncolytics 2019, 12, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli Yaraki, M.; Karami Tehrani, F. Apoptosis Induced by 13-S-hydroxyoctadecadienoic acid in the Breast Cancer Cell Lines, MCF-7 and MDA-MB-231. Iran. J. Basic Med. Sci. 2013, 16, 661–669. [Google Scholar] [PubMed]
- Shureiqi, I.; Jiang, W.; Zuo, X.; Wu, Y.; Stimmel, J.B.; Leesnitzer, L.M.; Morris, J.S.; Fan, H.Z.; Fischer, S.M.; Lippman, S.M. The 15-lipoxygenase-1 product 13-S-hydroxyoctadecadienoic acid down-regulates PPAR-delta to induce apoptosis in colorectal cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 9968–9973. [Google Scholar] [CrossRef] [PubMed]
- Yuri, M.; Sasahira, T.; Nakai, K.; Ishimaru, S.; Ohmori, H.; Kuniyasu, H. Reversal of expression of 15-lipoxygenase-1 to cyclooxygenase-2 is associated with development of colonic cancer. Histopathology 2007, 51, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; Shen, C.C.; Yi, Y.C.; Tsai, J.J.; Wang, C.C.; Chueh, J.T.; Lin, K.L.; Lee, T.C.; Pan, H.C.; Sheu, M.L. Honokiol inhibits gastric tumourigenesis by activation of 15-lipoxygenase-1 and consequent inhibition of peroxisome proliferator-activated receptor-gamma and COX-2-dependent signals. Br. J. Pharmacol. 2010, 160, 1963–1972. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Xia, H.H.; Tu, S.P.; Fan, D.M.; Lin, M.C.; Kung, H.F.; Lam, S.K.; Wong, B.C. 15-Lipoxygenase-1 mediates cyclooxygenase-2 inhibitor-induced apoptosis in gastric cancer. Carcinogenesis 2003, 24, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Dai, H.; Chen, S.; Zhang, Y.; Wu, T.; Cao, X.; Zhao, G.; Xu, A.; Wang, J.; Wu, L. Disruption of Chromosomal Architecture of cox2 Locus Sensitizes Lung Cancer Cells to Radiotherapy. Mol. Ther. J. Am. Soc. Gene Ther. 2018, 26, 2456–2465. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Tan, Q.; Luo, Q. Decreased expression level and DNA-binding activity of specificity protein 1 via cyclooxygenase-2 inhibition antagonizes radiation resistance, cell migration and invasion in radiation-resistant lung cancer cells. Oncol. Lett. 2018, 16, 3029–3037. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.Q.; Liao, H.; Shi, F.; Chen, X.P.; Hu, H.C.; Tian, M.Q.; Wang, L.H.; Ying, S. Inhibition of cyclooxygenase-2 sensitizes lung cancer cells to radiation-induced apoptosis. Oncol. Lett. 2017, 14, 5959–5965. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Mao, F.; Zuo, X.; Moussalli, M.J.; Elias, E.; Xu, W.; Shureiqi, I. 15-LOX-1 suppression of hypoxia-induced metastatic phenotype and HIF-1alpha expression in human colon cancer cells. Cancer Med. 2014, 3, 472–484. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Wortmann, M.; Mandal, P.K.; Arpornchayanon, W.; Jannasch, K.; Alves, F.; Strieth, S.; Conrad, M.; Beck, H. Absence of glutathione peroxidase 4 affects tumor angiogenesis through increased 12/15-lipoxygenase activity. Neoplasia 2010, 12, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Moreno Roig, E.; Groot, A.J.; Yaromina, A.; Hendrickx, T.C.; Barbeau, L.M.O.; Giuranno, L.; Dams, G.; Ient, J.; Olivo Pimentel, V.; van Gisbergen, M.W.; et al. HIF-1alpha and HIF-2alpha Differently Regulate the Radiation Sensitivity of NSCLC Cells. Cells 2019, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wu, L.; Li, D.; Xu, Y.; Zhang, L.; Niu, K.; Kong, R.; Gu, J.; Xu, Z.; Chen, Z.; et al. Radiosensitizing effects of miR-18a-5p on lung cancer stem-like cells via downregulating both ATM and HIF-1alpha. Cancer Med. 2018, 7, 3834–3847. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.W.; Elledge, S.J. The DNA damage response: Ten years after. Mol. Cell 2007, 28, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Niida, H.; Nakanishi, M. DNA damage checkpoints in mammals. Mutagenesis 2006, 21, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; de Toledo, S.M.; Pandey, B.N.; Guo, G.; Pain, D.; Li, H.; Azzam, E.I. Role of the translationally controlled tumor protein in DNA damage sensing and repair. Proc. Natl. Acad. Sci. USA 2012, 109, E926–E933. [Google Scholar] [CrossRef] [PubMed]
- Yoshiyama, K.O.; Sakaguchi, K.; Kimura, S. DNA damage response in plants: Conserved and variable response compared to animals. Biology 2013, 2, 1338–1356. [Google Scholar] [CrossRef] [PubMed]
- Huhn, D.; Bolck, H.A.; Sartori, A.A. Targeting DNA double-strand break signalling and repair: Recent advances in cancer therapy. Swiss Med. Wkly. 2013, 143, w13837. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Feng, Y.; Schultz, C.J.; Li, X.A.; Wu, H.; Wang, D. Synergistic effect of 15-lipoxygenase 2 and radiation in killing head-and-neck cancer. Cancer Gene Ther. 2008, 15, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Lovey, J.; Nie, D.; Tovari, J.; Kenessey, I.; Timar, J.; Kandouz, M.; Honn, K.V. Radiosensitivity of human prostate cancer cells can be modulated by inhibition of 12-lipoxygenase. Cancer Lett. 2013, 335, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Youn, H.; Seong, K.M.; Yun, Y.J.; Kim, W.; Kim, Y.H.; Lee, J.Y.; Kim, C.S.; Jin, Y.W.; Youn, B. Psoralidin, a dual inhibitor of COX-2 and 5-LOX, regulates ionizing radiation (IR)-induced pulmonary inflammation. Biochem. Pharmacol. 2011, 82, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Middleton, F.K.; Pollard, J.R.; Curtin, N.J. The Impact of p53 Dysfunction in ATR Inhibitor Cytotoxicity and Chemo- and Radiosensitisation. Cancers 2018, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.F.; Sun, W.Y.; Yu, D.H.; Zhao, Y.; Xu, H.M.; He, Y.F.; Li, H.J. Rotundic acid enhances the impact of radiological toxicity on MCF-7 cells through the ATM/p53 pathway. Int. J. Oncol. 2018, 53, 2269–2277. [Google Scholar] [CrossRef] [PubMed]
- Szoltysek, K.; Janus, P.; Zajac, G.; Stokowy, T.; Walaszczyk, A.; Widlak, W.; Wojtas, B.; Gielniewski, B.; Cockell, S.; Perkins, N.D.; et al. RRAD, IL4I1, CDKN1A, and SERPINE1 genes are potentially co-regulated by NF-kappaB and p53 transcription factors in cells exposed to high doses of ionizing radiation. BMC Genom. 2018, 19, 813. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xiong, Y.; Ding, X.; Wang, L.; Zhao, Y.; Fei, Y.; Zhu, Y.; Shen, X.; Tan, C.; Liang, Z. Cathepsin L activated by mutant p53 and Egr-1 promotes ionizing radiation-induced EMT in human NSCLC. J. Exp. Clin. Cancer Res. 2019, 38, 61. [Google Scholar] [CrossRef] [PubMed]
- Costanzi, C.; Pehrson, J.R. MACROH2A2, a new member of the MARCOH2A core histone family. J. Biol. Chem. 2001, 276, 21776–21784. [Google Scholar] [CrossRef] [PubMed]
- Tanasijevic, B.; Rasmussen, T.P. X chromosome inactivation and differentiation occur readily in ES cells doubly-deficient for macroH2A1 and macroH2A2. PLoS ONE 2011, 6, e21512. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wang, Y.; Yang, Q.; Li, X.; Kong, X.; Zhang, N.; Yuan, C.; Yang, N.; Kong, B. Low-dose radiation-induced epithelial-mesenchymal transition through NF-kappaB in cervical cancer cells. Int. J. Oncol. 2013, 42, 1801–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamoto, A.; Yokoe, T.; Tanaka, K.; Saigusa, S.; Toiyama, Y.; Yasuda, H.; Inoue, Y.; Miki, C.; Kusunoki, M. Radiation induces epithelial-mesenchymal transition in colorectal cancer cells. Oncol. Rep. 2012, 27, 51–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Na, Y.J.; Kim, B.R.; Kim, J.L.; Kang, S.; Jeong, Y.A.; Park, S.H.; Jo, M.J.; Kim, J.-Y.; Kim, H.J.; Oh, S.C.; et al. Deficiency of 15-LOX-1 Induces Radioresistance through Downregulation of MacroH2A2 in Colorectal Cancer. Cancers 2019, 11, 1776. https://doi.org/10.3390/cancers11111776
Na YJ, Kim BR, Kim JL, Kang S, Jeong YA, Park SH, Jo MJ, Kim J-Y, Kim HJ, Oh SC, et al. Deficiency of 15-LOX-1 Induces Radioresistance through Downregulation of MacroH2A2 in Colorectal Cancer. Cancers. 2019; 11(11):1776. https://doi.org/10.3390/cancers11111776
Chicago/Turabian StyleNa, Yoo Jin, Bo Ram Kim, Jung Lim Kim, Sanghee Kang, Yoon A. Jeong, Seong Hye Park, Min Jee Jo, Jeong-Yub Kim, Hong Jun Kim, Sang Cheul Oh, and et al. 2019. "Deficiency of 15-LOX-1 Induces Radioresistance through Downregulation of MacroH2A2 in Colorectal Cancer" Cancers 11, no. 11: 1776. https://doi.org/10.3390/cancers11111776